

Research Article: Journal of Drug and Alcohol Research (2024) Volume 13, Issue 10
Genetically Predicted Omega-3 Fatty Acids Mediate the Association between SM23-33 Abundance in Stool and Childhood Obesity: A Mendelian Randomization Study.
Min Zhang1, Jinke Li2, Fang Huang1 and Wenjuan Yan3*2Department of Family Medicine, Datuan Community Health Service Center, China
3Department of Medical Functional Examination, Guangming Hospital of Traditional Chinese Medicine, China
Wenjuan Yan, Department of Medical Functional Examination, Guangming Hospital of Traditional Chinese Medicine, China, Email: zhm395010878@gmail.com
Received: 02-Oct-2024, Manuscript No. JDAR-24-149796 ; Editor assigned: 04-Oct-2024, Pre QC No. JDAR-24-149796 (PQ); Reviewed: 18-Oct-2024, QC No. JDAR-24-149796; Revised: 23-Oct-2024, Manuscript No. JDAR-24-149796 (PQ); Published: 30-Oct-2024, DOI: 10.4303/JDAR/236411
Abstract
Background: Childhood obesity may be linked to gut microbiota and omega-3 fatty acids, but the precise relationships remain unclear.
Objective: This study aimed to investigate the relationships between omega-3 fatty acids, the abundance of SM23-33 in stool, and childhood
obesity.
Methods: We performed Linkage Disequilibrium Score Regression (LDSC) to assess the genetic correlations among the abundance of SM23-33 in stool, omega-3 fatty acids, and childhood obesity. Additionally, bidirectional Mendelian Randomization (MR) analyses were conducted to explore the causal connections between SM23-33 abundance in stool and childhood obesity. A two-step MR analysis was used to determine whether omega-3 fatty acids, specifically Docosahexaenoic Acid (DHA), mediated these relationships. All statistical analyses were conducted using R software, adhering to the STROBE-MR checklist for reporting MR studies.
Results: No significant genetic correlation was found among SM23-33 abundance in stool, omega-3 fatty acids, and childhood obesity (rg_p>0.05). However, MR analysis indicated a causal association between SM23-33 abundance in stool and childhood obesity (OR=0.747, 95% CI: 0.584-0.957, P=0.021). There was no strong evidence that genetically predicted childhood obesity affected SM23-33 abundance in stool. The two-step MR analysis suggested that the association between SM23-33 abundance in stool and childhood obesity was mediated by DHA, with a mediation proportion of 3.56% (95% CI: 3.43%, 3.69%).
Conclusion: This study provides evidence supporting a causal relationship between the abundance of SM23-33 in stool and childhood obesity, with a potential mediating effect by Docosahexaenoic Acid (DHA).
Keywords
SM23-33 abundance in stool; Omega-3 fatty acids; Mendelian randomization; Docosahexaenoic acid; Childhood obesity
Introduction
Obesity is a complex, chronic condition characterized by excessive fat accumulation that can negatively affect health. It increases the risk of type 2 diabetes, cardiovascular diseases, bone health issues, reproductive problems, and certain cancers [1]. It also impacts quality of life, affecting sleep and mobility [2]. According to the World Health Organization, in 2022, one in 8 people worldwide were obese. Since 1990, the prevalence of obesity in adults has more than doubled, while the rate among adolescents has increased by 4 times. By 2022, over 390 million children and adolescents aged 5-19 were overweight, including 160 million with obesity. These alarming statistics underscore the urgent need for effective strategies to combat this global health crisis.
The gut microbiome significantly influences the development and progression of obesity [3,4]. Alterations in the gut microbiome are associated with childhood obesity, marked by reduced diversity of beneficial bacteria and increased presence of opportunistic pathogens. Research shows that obese children have distinct gut microbiome profiles compared to lean children. For instance, the Firmicutes phylum is more abundant in obese children, while the Bacteroidetes phylum is more prevalent in lean children [5]. Additionally, the gut microbiome in obese children generally exhibits lower diversity and a higher tendency towards inflammation [6,7]. The gut microbiome regulates energy harvest and storage, and its alterations can change metabolic pathways, contributing to childhood obesity [7]. It also affects appetite and satiety hormones like ghrelin and leptin, potentially leading to overeating and weight gain in children [8]. Modulating the gut microbiome through dietary interventions, such as prebiotics or probiotics, has proven effective in managing weight in children [6].
Despite the established link between the gut microbiome and childhood obesity, much remains to be understood about the specific roles of different microbial families. The SM23-33 order, consisting of strictly fermentative Phycisphaerae bacteria, is one such group that requires further exploration. Comprising 2 families, SM23-33 and FEN-1343, these bacteria were identified in metagenomic datasets from estuary and sulfur-rich hydrothermal sediments [9]. Although literature on this order is limited, recent research suggests their functional capacity is more complex than previously known. These bacteria thrive in environments lacking electron acceptors and can degrade complex carbon substrates. Some members may perform anaerobic sulfite reduction, indicating their metabolic diversity and adaptability [10]. Although no direct evidence links SM23-33 bacteria to childhood obesity, their diverse metabolic abilities and roles in complex carbon metabolism suggest further research could uncover connections to childhood obesity and other complex diseases.
While investigating potential links between SM23- 33 bacteria and childhood obesity, other studies have examined the role of nutrition, particularly omega-3 fatty acids, in preventing and managing this condition. Omega-3 fatty acids, including Alpha-linolenic Acid (ALA), Docosahexaenoic Acid (DHA), Docosapentaenoic Acid (DPA), and Eicosapentaenoic Acid (EPA), may help combat childhood obesity by reducing inflammation, enhancing insulin sensitivity, and improving cardiovascular health [11-13]. They might also affect appetite control and satiety, potentially resulting in lower energy intake and improved weight management [14]. A systematic review conducted in 2019 revealed that omega-3 supplementation led to notable reductions in BMI and waist circumference among children and adolescents [15]. A 2023 study reported improved insulin sensitivity and reduced inflammatory markers in obese children receiving omega-3 supplements [16]. However, some studies have yielded inconclusive or contradictory results. For example, a 2018 study noted an increase in BMI and body fat percentage in obese children following omega-3 supplementation [17]. A 2021 review of 20 clinical trials found no significant effect of omega-3 supplementation on BMI or body fat percentage in children [18]. These mixed findings emphasize the importance of further investigation to better understand the links between omega-3 fatty acids, gut microbiota, and childhood obesity.
Mendelian Randomization (MR) analysis is a powerful epidemiological tool based on Mendelian inheritance principles. This technique uses genetic variants as instrumental variables to estimate causal relationships, minimizing confounding variables, measurement errors, and reverse causation [19]. Based on recent studies, we hypothesize a causal association between SM23-33 abundance in stool and childhood obesity, with omega-3 fatty acids acting as mediators. We utilized mediation MR analysis to explore this relationship, employing genetic data from extensive Genome-wide Association Studies (GWAS). Our aim is to clarify the causal mechanisms at play, offering a better understanding of how SM23-33 affects childhood obesity by influencing omega-3 profiles.
Materials and Methods
Study design
In this study, we utilized various Mendelian Randomization (MR) techniques, including genetic correlation analysis (LDSC), Two-sample MR (TSMR), Bidirectional MR (BDMR), multivariable MR (MVMR), and two-step MR (2SMR), to explore the causal relationships between the abundance of SM23-33 in stool, omega-3 fatty acids, and childhood obesity. Our study was grounded in 3 fundamental assumptions necessary for accurate MR estimates:
(i) Genetic Instrumental Variables (ivs) must be strongly associated with the exposure,
(ii) Genetic ivs must not be related to confounders affecting both the exposure and outcome, and
(iii) Genetic ivs should impact the outcome solely through their effect on the exposure [20].
By applying a range of MR methods, we aimed to reduce bias and produce reliable estimates of how modifiable exposures relate to the outcomes of interest. Additionally, we adhered to the STROBE-MR checklist to ensure the integrity of this observational MR study. Here is a detailed and transparent summary of our study design plan.
Step 1: GWAS summary data
• SM23-33 abundance in stool (Exposure)
• 5,959 Finnish individuals, FINRISK 2022, Qin Y et al. (2022)
• Childhood obesity (Outcomes)
• 13005 European individuals, EGG consortium, Bradfield JP et al. (2019)
• Omega-3 fatty acids (Mediator)
• 136,016 individuals from ALSPAC et al, Karjalainen MK et al. (2024)
• 8,299 individuals from CLSA et al, Chen Y et al. (2023)
Step 2: Data quality control
• Extracting European population data
• Genetic correlation analysis (LDSC regression)
Step 3: Instrumental variables
• Single nucleotide polymorphism
Step 4: Selection of genetic IVs
• Association with SM23-33 abundance in stool (P<5 × 10-5), omega-3 fatty acids (P<5 × 10-8), childhood obesity (P<5 × 10-5)
• LD R2<0.001, window size=10 kb
• Exclude the ambiguous SNPs with a minor allele frequency<0.01
• Remove potential pleiotropic SNPs by MR-PRESSO
• Remove SNPs associated with confounder
Step 5: Mendelian randomization
• Inverse variance weighted
• MR Egger
• Weighted median
• Simple mode
• Weighted mode
Step 6: Sensitivity analysis
• Cochran’s Q test (Heterogeneity)
• F statistic (Strength of IVs)
• R2 (The proportion of variance)
• MR-Egger intercept (Pleiotropic test)
• MR-PRESSO (Outlier test)
• Leave-one-out
Data sources
Source of SM23-33 abundance in stool: The GWAS data on SM23-33 abundance in stool used in this study were obtained from research investigating the genetic traits of gut microbiota. The original GWAS included 2,801 microbial taxa and 7,967,866 human genetic variants from 5,959 individuals in the FR02 cohort. The summary data for SM23-33 stool abundance was accessed via the GWAS Catalog (https://www.ebi.ac.uk/gwas), with the accession number GCST90032559. Further details about this GWAS can be found in the original study [21].
Data on DHA were obtained from the GWAS Catalog, originally compiled by Karjalainen MK et al. (2024), under accession number GCST90301955 [22]. This study investigated genetic associations with 233 circulating metabolic traits across 33 cohorts, involving a total of 136,016 participants, primarily of European descent, with 11.60% from Asia (15,775 out of 136,016). Data on other omega-3 fatty acids, including ALA, DPA, and EPA, were also retrieved from the GWAS Catalog, originally published by Chen Y et al., with accession numbers GCST90199748, GCST90199713, and GCST90200349, respectively [23]. This research was based on the Canadian Longitudinal Study on Aging (CLSA) cohort and analyzed 1,091 blood metabolites and 309 metabolite ratios from 8,299 participants and approximately 15.4 million SNPs. Detailed information on this GWAS can be found in the original publication.
Childhood obesity data were supplied by the EGG consortium and obtained from (www.egg-consortium. org) [24]. To identify additional genetic variants linked to childhood obesity, a trans-ancestral meta-analysis of 30 studies was conducted, including up to 13,005 cases and 15,599 controls from European, African, North/South American, and East Asian ancestries. European-specific GWAS data were subsequently extracted for this analysis.
Selection of IVs and data harmonization
To meet the rigorous criteria based on the 3 core Mendelian Randomization (MR) assumptions and to minimize horizontal pleiotropy, we selected only independent genome-wide significant Single Nucleotide Polymorphisms (SNPs) as instrumental variables (IVs) for the exposure. The chosen IVs had to be strongly associated with the exposure (SM23-33 abundance in stool, omega-3 fatty acids, childhood obesity), with SNPs significantly linked to the outcomes being selected at the genome-wide level (P<5×10−8, r2<0.001, window size=10 kb). In cases where too few SNPs met these criteria, the threshold was adjusted to (P<5 × 10−5 or P<1 × 10−5, r2<0.001, window size=10 kb). Additionally, we calculated the F statistics for the IVs to evaluate the risk of weak instrument bias. To minimize this bias, we retained working variables with F>10, and for datasets with a larger number of SNPs, only those with F>100 were included. The formula for calculating the F value and R2 is as follows.
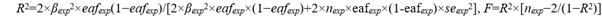
To prevent multicollinearity and confounding effects, we used the LD link tool (https://ldlink.nih.gov/?tab=ldtrait) to perform Linkage Disequilibrium (LD) pruning on the SNPs. Using the LD trait module, we focused on the European population with the CRCH38 genome build, applying parameters of r2=0.001 and a window size of 10,000 kb. This method ensured that only one representative SNP per window was retained, minimizing confounding from LD. We also excluded SNPs associated with various phenotypes, including BMI, weight, body mass index, obesity-related traits, whole body fat mass, waist-to-hip ratio adjusted for BMI, waist circumference adjusted for BMI, abdominal fat cell number, hip circumference adjusted for BMI, whole body fat-free mass, triglycerides, LDL, HDL, and VLDL, to further reduce potential confounding. This exclusion aimed to strengthen the reliability of our findings. After filtering the data, we conducted a horizontal pleiotropy analysis. If horizontal pleiotropy was detected, the MR-PRESSO test was performed, and any outliers identified by this test were removed before proceeding with the subsequent analysis.
Causal effects of SM23-33 abundance in stool on childhood obesity
We conducted bidirectional Mendelian Randomization (MR) analyses to explore the causal relationship between SM23-33 abundance in stool and childhood obesity, estimating the total effect (β) of this association. To estimate effects, we employed the inverse variance weighted (IVW) method, reporting β ± SE for continuous outcomes and Odds Ratios (OR) with 95% Confidence Intervals (CI) for binary outcomes. The IVW method involved meta-analyzing SNP-specific Wald estimates by dividing the SNP-outcome association by the SNPexposure association, using random effects to derive the final causal effect estimate. Additionally, we used MREgger and weighted median methods as complementary approaches to the IVW, offering a more comprehensive view of the causal relationship.
Mediation analyses connect ‘SM23-33 abundance in stool,’ ‘Omega-3 fatty acids,’ and ‘childhood obesity
We conducted a mediation analysis using a two-step Mendelian Randomization (MR) approach to investigate whether omega-3 fatty acids mediate the causal relationship between SM23-33 abundance in stool and childhood obesity [25]. The analysis involved 2 steps:
1. A uni-variable MR (UVMR) model was used to estimate the effect of the exposure on the mediator (omega-3 fatty acids), and
2. The multivariable MR (MVMR) model assessed the impact of the mediator on childhood obesity.
In the MVMR model, genetic variants for both the mediator and the exposure were included in both stages of regression to ensure that the mediator’s effect on the outcome was independent of the exposure, and to estimate the direct effect of the exposure on the outcome. The indirect effect was estimated by multiplying the regression estimates from the second stage.
For our study, the overall effect of SM23-33 abundance in stool on childhood obesity was decomposed into direct and indirect effects. The direct effect represents the impact of SM23-33 abundance in stool on childhood obesity without mediation, while the indirect effect represents the mediation effect through omega-3 fatty acids. The mediation effect was determined as β1 × β2, where β1 represents the causal impact of SM23-33 abundance in stool on Omega-3 fatty acids, and β2 reflects the causal influence of Omega-3 fatty acids on childhood obesity, adjusted for confounding factors related to the exposure. The percentage of mediation was determined by dividing the indirect effect by the total effect. Confidence intervals were calculated using the Delta method [26-32].
Sensitivity analysis
We assessed the directional association between each identified SNP and both the exposure and outcome variables using MR Steiger filtering. This method assesses the extent to which variation in the exposure and outcome can be attributed to instrumental SNPs and determines whether the variability in the outcome is less than that in the exposure. Horizontal pleiotropy was further examined using the MREgger approach, which involves weighted linear regression with an unconstrained intercept. This intercept serves as an indicator of the average pleiotropic effect across genetic variants, reflecting the typical direct influence of a variant on the outcome variable. A significant deviation of the intercept from zero (MR-Egger intercept P<0.05) suggests the presence of horizontal pleiotropy. Additionally, we used Cochrane’s Q-test to evaluate heterogeneity, with lower p-values indicating increased heterogeneity and a higher likelihood of directional pleiotropy. To identify potential SNP outliers, leave-one-out analyses were also conducted.
Statistical analysis
We used the results from the Inverse Variance Weighted (IVW) method as our primary analysis and applied a significance threshold of P<0.05 to the MR analysis, considering P-values below this threshold as statistically significant. All statistical analyses and data visualizations were conducted using R software (R Foundation, Vienna, Austria). Specifically, we used the TwoSampleMR package (https://github.com/MRCIEU/TwoSampleMR) for univariable and multivariable MR analyses, the GenomicSEM package (Yen-Tsung Huang, Patrick J. Smith, USA) for Linkage Disequilibrium Score Regression (LDSC) analysis, and the PNG package (Boutell, Netherlands) for data visualization.
LDSC was employed to estimate the genetic correlation (rg) between SM23-33 abundance in stool, omega-3 fatty acids, and childhood obesity. LDSC evaluates the relationship between test statistics and linkage disequilibrium to quantify the contribution of genuine polygenic signals or potential biases. This method allows for the assessment of genetic correlations using GWAS summary statistics while avoiding bias from sample overlap. We calculated the z-scores for each variant of Trait 1 and multiplied them by those of Trait 2. The genetic covariance was then obtained by regressing this product against the LD score. The genetic correlation was derived by normalizing the genetic covariance by SNP heritability, with statistical significance set at P<0.05.
Results
Selection of IVs
We screened the remaining 73 SNPs related to SM23- 33 abundance in stool and 120 SNPs related to omega-3 fatty acids that met assumptions 1, 2, and 3. We excluded SNPs associated with obesity-specifically rs147608546, rs1995755, rs41264899, rs75596315, rs7581869, rs7791602, rs79131883, and rs9783388-as well as SNP rs56154844 associated with omega-3 fatty acids. These exclusions were made based on prior evidence suggesting that these SNPs could act as confounders, potentially affecting the causal relationship between SM23-33 abundance in stool, omega-3 fatty acids, and childhood obesity. All Instrumental Variables (IVs) had F-statistics above 10, indicating no weak instrument bias. Specifically, the average F-value for the association between exposure and outcome was 18.85, for exposure and mediator it was 18.84, and for mediator and outcome it was 162.73. These values indicate that the instrumental variables used in the analysis are robust and dependable.
LDSC regression analysis
We conducted LDSC regression analysis to assess the genetic correlation among SM23-33 abundance in stool, omega-3 fatty acids, and childhood obesity. The results of this analysis indicated no significant genetic correlation among the 3 traits (rg_p>0.05). This finding supports the validity of our subsequent MR analysis, as detailed in Table 1.
Table 1: Genetic correlations among SM23-33 abundance in stool, Omega-3 fatty acids, and childhood obesity
| Trait1 | Trait2 | Rg | Rg_se | Rg_p |
|---|---|---|---|---|
| SM23-33 abundance in stool | Childhood obesity | 0.043 | 0.233 | 0.857 |
| SM23-33 abundance in stool | Alpha-linolenic acid | 1.281 | 0.676 | 0.058 |
| SM23-33 abundance in stool | Docosahexaenoic acid | 0.254 | 0.172 | 0.143 |
| SM23-33 abundance in stool | Docosapentaenoic acid | 0.613 | 0.588 | 0.298 |
| SM23-33 abundance in stool | Eicosapentaenoic acid | 0.065 | 0.63 | 0.918 |
| Alpha-linolenic acid | Childhood obesity | 0.144 | 0.155 | 0.361 |
| Docosahexaenoic acid | Childhood obesity | -0.026 | 0.048 | 0.597 |
| Docosapentaenoic acid | Childhood obesity | 0.131 | 0.136 | 0.337 |
| Eicosapentaenoic acid | Childhood obesity | 0.062 | 0.138 | 0.662 |
Causal association between SM23-33 abundance in stool with childhood obesity
We used 62 SNPs to genetically proxy the effect of SM23- 33 abundance in stool (Table S1). Our analysis revealed a negative correlation between SM23-33 abundance in stool and the risk of childhood obesity (Figure 1). Using inverse-variance weighted methods, we found an odds ratio (OR) of 0.747 (95% CI: 0.584-0.957; P=0.021) for childhood obesity per 1 standard deviation increase in SM23-33 abundance in stool. This implies that each standard deviation increase in SM23-33 abundance in stool is associated with a 25.3% reduction in the relative odds of childhood obesity. For further details, refer to the supplementary materials (Tables S2 and S3).

Figure 1: Univariable MR analysis shows causal effects of genetically proxied SM23-33 abundance in Stool, Omega-3 fatty Acids, and childhood obesity using IVW methods
Abbreviations: SNPs, single-nucleotide polymorphisms; OR, odds ratio; CI, confidence interval; SM23-33, SM23-33 abundance in stool; Obesity, childhood obesity; MR, Mendelian randomization; ALA, Alpha-linolenic Acid, DHA, Docosahexaenoic Acid; DPA, Docosapentaenoic Acid
Mediation MR analyses of Omega-3 fatty acids
Genetically proxied SM23-33 abundance in stool was significantly associated with childhood obesity [OR=0.747 (95% CI: 0.584-0.957), P=0.021]. To explore the mechanisms through which SM23-33 abundance in stool might influence childhood obesity, we examined its effect on several common omega-3 fatty acids. SM23-33 abundance in stool was significantly associated with DHA [OR=1.059 (95% CI: 1.012-1.108), P=0.012], but not with ALA [OR=1.014 (95% CI: 0.855-1.201), P=0.877], DPA [OR=1.057 (95% CI: 0.877-1.275), P=0.559], or EPA [OR=1.043 (95% CI: 0.872-1.248), P=0.646]. Similarly, genetically proxied ALA [OR=1.101 (95% CI: 0.974- 1.244), P=0.124], DPA [OR=1.022 (95% CI: 0.902-1.159), P=0.729], and EPA [OR=0.928 (95% CI: 0.850-1.013), P=0.095] were not significantly associated with childhood obesity. However, DHA remained significantly associated with childhood obesity [OR=0.835 (95% CI: 0.735-0.948), P=0.005], as shown in Figure 2.

Figure 2: Schematic diagram of the mediating effect of Omega-3 fatty acids between SM23-33 abundance in Stool and SM23-33 abundance in Stool. The solid line represents that the causal relationship is established, and the dotted line represents that the causal relationship is not established
To identify mediators in the causal relationship between SM23-33 abundance in stool and childhood obesity, we used the following criteria: A mediator must exhibit a direct causal effect from SM23-33 abundance in stool and have a direct causal effect on childhood obesity, with consistent directional relationships. Among the 4 omega-3 fatty acids analyzed (ALA, DHA, DPA, EPA), only DHA met these criteria as a mediator. Specifically, SM23-33 abundance in stool was positively associated with DHA [OR=1.059 (95% CI: 1.012-1.108), P=0.012], and DHA showed a negative association with childhood obesity [OR=0.831 (95% CI: 0.719-0.961), P=0.012]. Mediation analysis revealed that DHA mediated the effect of SM23-33 abundance in stool on childhood obesity, with a mediation effect of -0.010 (95% CI: -0.024 to -0.001), accounting for 3.56% (95% CI: 3.43% to 3.69%) of the total effect. For more details, see the supplementary materials (Table S4).
Reverse MR analysis
Initially, we investigated the causal relationship by using childhood obesity and omega-3 fatty acids as exposures, with SM23-33 abundance in stool as the outcome. This analysis sought to assess whether childhood obesity and omega-3 fatty acids have a causal impact on SM23-33 abundance in stool. Second, we reversed the roles by using childhood obesity as the exposure and omega-3 fatty acids as the outcome, to investigate whether childhood obesity causally influences omega-3 fatty acids.
The MR analysis revealed no evidence of reverse causation between SM23-33 abundance in stool, omega-3 fatty acids, and childhood obesity. Specifically, the results did not support the hypothesis that childhood obesity causes changes in SM23-33 abundance in stool or omega-3 fatty acids, nor did they suggest that omega-3 fatty acids affect SM23-33 abundance in stool. The MR estimates and results from the sensitivity analysis are presented in the supplementary material (Table S3).
Sensitivity analysis
To address potential pleiotropy in our causal effect estimates, we conducted several sensitivity analyses (Table 2). Cochran’s Q-test and funnel plot analysis showed no evidence of heterogeneity or asymmetry among the SNPs involved in the causal relationship. Furthermore, leaveone- out analysis verified that each SNP had a substantial impact on the overall causal estimates. Re-analysis of the MR study, after excluding individual SNPs, consistently produced similar results, indicating that all SNPs played a crucial role in establishing the causal relationship. Further details are available in the supplementary material (Tables S1 and S5).
Table 2: The results of the heterogeneity and horizontal pleiotropy
| Exposure | Outcome | MR Egger pleiotropy test | IVW Cochran’s Q text | MR-PRESSO | |||
|---|---|---|---|---|---|---|---|
| Intercept | SE | P_value | Q_test | P_value | outliers | ||
| SM23-33 | Obesity | -0.001 | 0.011 | 0.889 | 53.897 | 0.73 | No |
| SM23-34 | ALA | 0.002 | 0.006 | 0.599 | 77.592 | 0.223 | No |
| SM23-35 | DHA | -0.001 | 0.001 | 0.334 | 67.411 | 0.531 | No |
| SM23-36 | DPA | 0.002 | 0.008 | 0.77 | 93.856 | 0.026 | No |
| SM23-37 | EPA | 0.001 | 0.008 | 0.936 | 85.089 | 0.093 | No |
| ALA | Obesity | -0.005 | 0.017 | 0.807 | 12.525 | 0.898 | No |
| DHA | Obesity | -0.003 | 0.007 | 0.829 | 51.986 | 0.04 | No |
| DPA | Obesity | -0.009 | 0.019 | 0.665 | 31.685 | 0.046 | No |
| EPA | Obesity | -0.002 | 0.011 | 0.863 | 26.045 | 0.406 | No |
| Abbreviations: SM23-33, SM23-33 abundance in stool; Obesity, childhood obesity; ALA, Alpha-linolenic Acid, DHA, Docosahexaenoic Acid; DPA, Docosapentaenoic Acid
|
|||||||
Discussion
In this study, we explored the causal associations between SM23-33 abundance in stool, omega-3 fatty acids, and childhood obesity using large-scale genetic data and Mendelian Randomization (MR) analysis. To our knowledge, this is the inaugural MR analysis exploring these relationships. Our rigorous inclusion criteria and sensitivity analyses revealed potential causal links between SM23-33 abundance in stool and childhood obesity, with DHA identified as a specific omega-3 fatty acid mediator in this pathway [33-36].
Previous research has highlighted significant differences in gut microbiota composition between children with obesity and those with a healthy weight. In cases of childhood obesity, bacteria associated with energy metabolism, such as Bacteroidetes and Clostridium, are typically less abundant, while bacteria linked to inflammation, such as Prevotella, are more prevalent. Currently, no research has specifically addressed the relationship between SM23-33 abundance in stool and childhood obesity. Our study suggests that SM23- 33 abundance in stool may play a crucial role in childhood obesity, providing a foundation for future research on this topic.
Previous research has indicated that omega-3 fatty acids can improve insulin sensitivity, influence fat cell metabolism and storage, reduce inflammation and metabolic syndrome risk, and regulate appetite, lipid metabolism, and immune function. These effects are thought to contribute to the prevention of childhood obesity. In our study, we examined 4 omega-3 fatty acids: ALA, DHA, DPA, and EPA. Our results revealed that only DHA significantly reduced the risk of childhood obesity (OR=0.747, 95%CI: 0.584-0.957; P=0.021) and served as a weak mediator in the relationship between SM23-33 abundance in stool and childhood obesity (3.56%, 95% CI: 3.43%-3.69%). This suggests that DHA may be the key omega-3 fatty acid influencing childhood obesity.
DHA is known for its significant health benefits, including supporting nervous system development, enhancing cardiovascular health, regulating inflammation, boosting immune function, reducing oxidative stress, and managing blood lipids. If future research confirms DHA’s critical role in preventing and treating childhood obesity, it could offer an effective strategy for addressing this public health challenge and inform novel approaches for primary child healthcare providers to combat obesity.
The potential mechanisms by which SM23-33 abundance in stool may contribute to childhood obesity include:
• Gut-brain axis: SM23-33 abundance may influence brain function and behavior through the gut-brain axis, affecting appetite regulation and energy metabolism, which could impact obesity risk.
• Inflammation and metabolic syndrome: Chronic low-grade intestinal inflammation can lead to insulin resistance and metabolic syndrome. SM23-33 abundance might help mitigate this inflammation by protecting intestinal barrier function and preventing excessive immune activation, thereby reducing obesity risk.
• Energy metabolism: SM23-33 abundance in stool may affect childhood obesity by modulating energy metabolism. Certain intestinal bacteria, such as Bacteroides, can break down complex carbohydrates into short-chain fatty acids, promoting energy absorption and storage, while others like Bifidobacteria may reduce fat accumulation by inhibiting energy absorption.
• Hormone secretion: SM23-33 abundance in stool may influence obesity by secreting hormones that regulate metabolism and appetite. For instance, some bacteria produce leptin and leptin-like peptides, which are involved in appetite and energy regulation.
• Metabolite production: SM23-33 abundance in stool could affect obesity development through the production of metabolites by specific microorganisms, such as short-chain fatty acids and bile acids, which influence host metabolism and energy regulation.
The potential significance of SM23-33 abundance in stool for preventing and treating childhood obesity includes:
Probiotics and prebiotics: Probiotics are live microorganisms that provide health benefits when consumed in sufficient quantities. Prebiotics, such as inulin and fructans, are indigestible substances that support the growth of beneficial intestinal bacteria. The production of probiotics and prebiotics associated with SM23-33 can influence gut microbiota composition and improve gut health, potentially helping to prevent childhood obesity.
Fecal flora transplantation: This approach involves transferring fecal microbiota from healthy donors to recipients to restore a balanced microbiome. Fecal flora transplantation has shown promise in obesity treatment, and SM23-33 could emerge as a new target for effective therapeutic interventions.
Understanding the link between gut microbiota and childhood obesity is complex and requires further research. Future research should aim to elucidate the precise mechanisms through which gut microbiota contribute to obesity in children and develop effective interventions.
Our study benefits from utilizing a large GWAS database and applying robust MR analysis methods. We employed various techniques, such as genetic correlation regression analysis (LDSC) and reverse MR analysis, to ensure the study’s rigor. However, there are limitations to consider:
Pleiotropy and heterogeneity: Despite using MR-PRESSO and MR-Egger methods to assess heterogeneity and pleiotropy, and the LD link website to remove potentially confounding SNPs, residual horizontal pleiotropy could still affect causal effect estimates.
GWAS database variation: The heterogeneity between different GWAS databases could impact the accuracy of our study’s results.
Ethnic diversity: Our study predominantly included individuals of European ancestry, with a small proportion from other ethnic groups. This could introduce slight bias and limit the applicability of our findings to other populations.
Influencing factors: Factors such as demographics, diet, and medication can affect gut microbiota and introduce variability, potentially reducing the statistical power and robustness of our results [37-39].
Conclusion
In summary, this study is the first to comprehensively examine the causal relationships between SM23-33 abundance in stool, Omega-3 fatty acids, and childhood obesity, highlighting DHA’s role. The findings provide valuable insights into childhood obesity’s etiology and suggest potential therapeutic and preventive strategies. We also explored possible mechanisms linking SM23- 33 abundance in stool to childhood obesity, offering new perspectives on gut microbiota-based treatments for this condition.
Ethical Approval
This research utilized publicly accessible data from GWAS studies. Each individual study included in the GWAS was approved by the respective Institutional Review Board, and participants or their authorized representatives provided informed consent.
Competing Interests
The author declares no competing interests related to this research.
Authors Contributions
Min Zhang conceived and designed the study, performed the data analysis and interpretation, and wrote the manuscript. Jinke Li and Wenjuan Yan were responsible for conceptualizing the study. Additionally, Fang Huang participated in the research of the related project.
Funding
This work was supported by Shanghai Pudong New District Health Commission Health Science and Technology Project (Grant numbers PW2021A-76).
Availability of Data and Materials
The analysis utilized publicly available datasets. Detailed information on all original contributions can be found in the article. All raw data can be downloaded from https:// ftp.ebi.ac.uk/pub/databases/gwas/summary_statistics/. For further inquiries, please contact the corresponding author.
Conflict Of Interest
The authors declare that they have no conflict of interest.
Acknowledgement
None.
References
- S. Pati, W. Irfan, A. Jameel, S. Ahmed, R.K. Shahid, Obesity and cancer: A current overview of epidemiology, pathogenesis, outcomes, and management, Cancers (Basel), 15(2023):485.
- B. Morrissey, E. Taveras, S. Allender, C. Strugnell, Sleep and obesity among children: A systematic review of multiple sleep dimensions, Pediatr Obes, 15(2020):e12619.
- Z. Cheng, L. Zhang, L. Yang, H. Chu, The critical role of gut microbiota in obesity, Front Endocrinol (Lausanne), 13(2022):1025706.
- Z. Zhuang, P. Zhou, J. Wang, X. Lu, Y. Chen, The characteristics, mechanisms and therapeutics: Exploring the role of gut microbiota in obesity, Diabetes Metab Syndr Obes, 16(2023):3691-3705.
- S. Zhang, Y. Dang, Roles of gut microbiota and metabolites in overweight and obesity of children, Front Endocrinol (Lausanne), 13(2022):994930.
- K.Y. Cho, Association of gut microbiota with obesity in children and adolescents, Clin Exp Pediatr, 66(2023):148-154.
- C.O. Akagbosu, E.P. Nadler, S. Levy, S.K. Hourigan, The role of the gut microbiome in pediatric obesity and bariatric surgery, Int J Mol Sci, 23(2022):15421.
- M. Petraroli, E. Castellone, V. Patianna, S. Esposito, Gut microbiota and obesity in adults and children: The state of the art, Front Pediatr, 9(2021):657020.
- B.J. Baker, C.S. Lazar, A.P. Teske, G.J. Dick, Genomic resolution of linkages in carbon, nitrogen, and sulfur cycling among widespread estuary sediment bacteria, Microbiome, 3(2015):14.
- W.B. Lenferink, T.A. van Alen, M.S.M. Jetten, H.J.M. Op den Camp, M.A.H.J. van Kessel, et al. Genomic analysis of the class Phycisphaerae reveals a versatile group of complex carbon-degrading bacteria, Antonie van Leeuwenhoek, 117(2023):2-5.
- S.C.R. Sherratt, R.P. Mason, P. Libby, P.G. Steg, D.L. Bhatt, Do patients benefit from omega-3 fatty acids? Cardiovasc Res, 119(2024):2884-2901.
- A. Borja-Magno, M. Guevara-Cruz, A. Flores-Lopez, S. Carrillo-Dominguez, J. Granados, Differential effects of high dose omega-3 fatty acids on metabolism and inflammation in patients with obesity: Eicosapentaenoic and docosahexaenoic acid supplementation, Front Nutr, 10(2023):1156995.
- S. Sinha, M. Haque, H. Lugova, S. Kumar, The effect of omega-3 fatty acids on insulin resistance, Life (Basel), 13(2023):1322.
- I.E. Young, H.M. Parker, R.L. Cook, N.J. O'dwyer, M.L. Garg, et al. Association between obesity and omega-3 status in healthy young women, Nutrients, 12(2020):1480.
- S. Jazayeri, J. Heshmati, Z. Mokhtari, M. Sepidarkish, G. Nameni, et al. Effect of omega-3 fatty acids supplementation on anthropometric indices in children and adolescents: A systematic review and meta-analysis of randomized controlled trials, Complement Ther Med, 53(2020):102487.
- B. Virgolici, L.A. Popescu, H.M. Virgolici, C.D. Stefan, M. Mohora, et al. Effects of omega-3 fatty acids associated with antioxidant vitamins in overweight and obese children, Acta Endocrinol (Buchar), 19(2023):221-227.
- V. Svensson, E. Johansson, M. Fischer, S.L. Deng, M. Hagstromer, et al. Omega-3 fatty acids does not affect physical activity and body weight in primary school children-a double-blind randomized placebo-controlled tria, Sci Rep, 8(2018):12725.
- F.M. Delpino, L.M. Figueiredo, B.G.C. da Silva, Effects of omega-3 supplementation on body weight and body fat mass: A systematic review, Clin Nutr, 44(2021):122-129.
- G. Hemani, J. Zheng, B. Elsworth, K.H. Wade, V. Haberland, et al. The MR-base platform supports systematic causal inference across the human phenome, Elife, 7(2018):e34408.
- D.A. Lawlor, R.M. Harbord, J.A.C. Sterne, N. Timpson, G. Davey Smith, Mendelian randomization: Using genes as instruments for making causal inferences in epidemiology, Stat Med, 27(2008):1133-63.
- Y. Qin, A.S. Havulinna, Y. Liu, P. Jousilahti, S.C. Ritchie, et al. Combined effects of host genetics and diet on human gut microbiota and incident disease in a single population cohort, Nat Genet, 54(2022):134-142.
- M.K. Karjalainen, S. Karthikeyan, C. Oliver-Williams, E. Sliz, E. Allara, et al. Genome-wide characterization of circulating metabolic biomarkers, Nature, 628(2024):130-138.
- Y. Chen, T. Lu, U. Pettersson-Kymmer, I.D. Stewart, G. Butler-Laporte, et al. Genomic atlas of the plasma metabolome prioritizes metabolites implicated in human diseases, Nat Genet, 55(2023):44-53.
- J.P. Bradfield, S. Vogelezang, J.F. Felix, A. Chesi, O, Helgeland, et al. A trans-ancestral meta-analysis of genome-wide association studies reveals loci associated with childhood obesity, Hum Mol Genet, 28(2019):3327-333.
- S. Burgess, R.M. Daniel, A.S. Butterworth, S.G. Thompson, Network mendelian randomization: Using genetic variants as instrumental variables to investigate mediation in causal pathways, Int J Epidemiol, 44(2022):484-95.
- M. Kalliomaki, M.C. Collado, S. Salminen, E. Isolauri, Early differences in fecal microbiota composition in children may predict overweight, Am J Clin Nutr, 87(2008):534-8.
- F. Borgo, E. Verduci, A. Riva, C. Lassandro, E. Riva, et al. Relative abundance in bacterial and fungal gut microbes in obese children: A case control study, Child Obes, 13(2017):78-84.
- G. Riccardi, R. Giacco, A.A. Rivellese, Dietary fat, insulin sensitivity and the metabolic syndrome, Clin Nutr, 23(2004):447-56.
- L. Lauritzen, P. Brambilla, A. Mazzocchi, L.B.S. Harslof, V. Ciappolino, DHA effects in brain development and function, Nutrients, 8(2016):6.
- S. Serini, E. Fasano, E. Piccioni, A.R.M. Cittadini, G. Calviello, Dietary n-3 polyunsaturated fatty acids and the paradox of their health benefits and potential harmful effects, Chem Res Toxicol, 24(2011):2093-105.
- T.A. Jacobson, S.B. Glickstein, J.D. Rowe, P.N. Soni, Effects of eicosapentaenoic acid and docosahexaenoic acid on low-density lipoprotein cholesterol and other lipids: A review, J Clin Lipidol, 6(2012):5-18.
- M. Valles-Colomer, G. Falony, Y. Darzi, E.F. Tigchelaar, J. Wang, et al. The neuroactive potential of the human gut microbiota in quality of life and depression, Nat Microbiol, 4(2019):623-632.
- P.D. Cani, A. Everard, Talking microbes: When gut bacteria interact with diet and host organs, Mol Nutr Food Res, 60(2016):58-66.
- A. Koh, F. De Vadder, P. Kovatcheva-Datchary, F. Backhed, From dietary fiber to host physiology: Short-chain fatty acids as key bacterial metabolites, Cell, 165(2016):1332-1345.
- Y.N. Yin, Q.F. Yu, N. Fu, X.W. Liu, F.G. Lu, Effects of four bifidobacteria on obesity in high-fat diet induced rats, World J Gastroenterol, 16(27):3394-401.
- K.E. Bouter, D.H. van Raalte, A.K. Groen, M. Nieuwdorp, Role of the gut microbiome in the pathogenesis of obesity and obesity-related metabolic dysfunction, Gastroenterology, 152(2107):1671-1678.
- A.C. Gomes, C. Hoffmann, J.F. Mota, The human gut microbiota: Metabolism and perspective in obesity, Gut Microbes, 9(2108):308-325.
- B. Sanchez, S. Delgado, A. Blanco-Miguez, A. Lourenco, M. Gueimonde, et al. Probiotics, gut microbiota, and their influence on host health and disease, Mol Nutr Food Res, 61(2107).
- X. Gu, Z.H. Chen, S.C. Zhang, Fecal microbiota transplantation in childhood: Past, present, and future, World J Pediatr, 19(2023):813-822.
Copyright: © 2024 Min Zhang, et al. This is an open access article distributed under the terms of the Creative Commons Attribution License, which permits unrestricted use, distribution, and reproduction in any medium, provided the original work is properly cited.